#Gene Cleaves
Explore tagged Tumblr posts
Visit Tumblr Blog
Explore Tumblr blogs with no restrictions, modern design and the best experience.
Last Seen Tumblr Blogs





Fun Fact
There were a total of 171.5 billion posts on Tumblr in 2019.
Text

BEFORE HE REALLY WENT "CUCKOO," SONNY WORE A PIN-STRIPED SUIT.
PIC INFO: Spotlight on Sonny the Cuckoo Bird in his earliest cereal box art incarnation, and was designed by Gene Cleaves in 1962. Sonny's first cereal box appearance was made in the mid '60s, a wholenseven years after Cocoa Puff's first hit market shelves.
Source: www.pinterest.com/pin/370913719307824920.
#Cukoo for Cocoa Puffs#Cocoa Puffs Cereal#Cocoa Puffs Breakfast Cereal#Cereal#Cuckoo for Cocoa Puffs#Cartoon Art#Cereal Box Art#Cereal Box#Box Art#Americana#1960s#Good Morning#General Mills Cereal#Breakfast Cereal#General Mills#Sonny the Cuckoo Bird#Breakfast#Gene Cleaves#Cocoa Puffs#Sixties#60s#American Style#1962#Vintage Kidcore
2 notes
·
View notes
Text
You want a baby. Simon can't get over his hangups to give you one. The solution to both problems? Johnny.
18+ SMUT. breeding. mildly dubious consent. Johnny feasts on your pussy and then does his best to knock you up while Simon watches. slight body worship. bastardization of religious imagery. Mean!Dom Simon. rough, messy sex.
He's not the type to saw off his own hand to feed you, but would rather find a third man to satiate you both. The only one who can care for you, he said. Can't do that when he's dead, can he?
Maybe that's why he calls for Johnny.
down boy. eager mutt. lil' pyedogs got himself all twisted up in a rutt. help him, won't you, pet?
Johnny's softer than Simon but only just. This margin of distance, however, could be the gaping maw of a canyon for how wide it really is when scaled down to fit. Boxed inside a narrow bed—on your belly, cheek on Simon's knee; ass up, legs spread. Johnny behind you—colluvium to Simon's mountainside, but still so broad, so thick, your hips twinge with the effort of keeping your knees so wide apart.
You feel it whistling through the chasm when he licks his lips behind you—a loud, lascivious smack, a wet suckle—and feel the burn of his stare riveted on the split of your flesh. This bare seam Simon swears he found nirvana tucked deep inside of. A buried ravine. Aquifer he quenches himself on.
A pilgrimage Johnny has been aching to take.
And that's what this is, isn't it? Yatra to the hidden piscina. A procession to pollute the tarn—something Simon can't bring himself to do.
Bad genes. Trauma—sticky, noxious tar that oozes from the rotting filaments; festering deep inside. Cancerous: a mass you long to cleave from bone but know it's not cosmetic. Not just the ball joints, or the studs, but the foundation itself. If you start tearing up pieces now you'll have nothing but an empty plot and a pile of damaged debris.
So:
Enter the third man.
A tool. Vassel. Pays fealty by fucking a baby into your womb.
It's what you wanted, isn't it?
(yes, but—)
It happens faster than you can keep up with. Hands on your hips. Coarse hair tickling the back of your thigh. Warm breath against sticky, wet flesh. A broad nose parting your folds. Inhale. Exhale on a deep, reedy groan.
"fuck, ye smell heavenly, doe."
Simon hums before you can peel your tongue from the roof of your mouth, answering for you with a brassy invitation: tastes even better, Johnny.
It's all the permission he needs before he pushes his head closer to your bare cunt, groaning as his tongue cleaves a silky, thick line between your folds. Gorging himself without much preamble. Hands curled around your hips like expensive silverware, pulling you back into the wanting, eager suck of his mouth.
All at once, it's too much. Your hips shift, squirming away from his tongue, the too-sharp press of his teeth against soft, sensitive flesh. Mewling, whimpering into the rain-wet fabric of Simon's jeans.
His hand falls on your head. A gentle tap. Behave, it says, but you can't.
Johnny tramples over that thin line between pleasure and ecstasy, blurring them both until it becomes pain. Overwhelming. Shoving you towards the edge before you've readied yourself for the fall.
"Can't, Simon, can't—"
The words elide, slurring into a high-pitched whine as Johnny feasts on your cunt. Devours you from the inside out—all teeth and tongue, sucking your clit until your thighs cramp from how tight your muscles tense, bleeding lactic acid over sore flesh. The scrape of his stubble over your folds, chafing them until they are raw. Swollen. Drenched hole fucked with the spear of his tongue, digging so deep you begin to fear that he's trying to crawl inside of you. Salt your womb with his own two hands—
"Can take it, birdie," is all Simon says before his hand slides down your arched, trembling spine. Fingers digging into the meat of your cheek, spreading you wider for Johnny to eat. "Look how eager he is. Can't get enough of that sweet cunt."
"It's—it's too much—"
You don't feel him move. Can't see much from the blurry tears in your eyes. But his other hand whips out, cracking over your untouched cheek in a firm, burning smack. One that makes Johnny moan when it lands. Cruel. Open palm. Hard enough to leave a welt in the shape of his hand—something that makes him groan when he sees it.
"fuckin' hell—" his fingers dig into the aching flesh, grip bruising.
Johnny peels his wet, open mouth away long enough to pant into the slick spread of your cunt, resting his cheek on the swell of your ass. "Bit rough wit' 'er, Lt."
Simon considers it. Body shaking the bed when he shrugs, leaning back to trail his hand back up your spine, curling over the arch of your nape. Keeping you still as you sob into his knee. "She likes it."
"know she does. Fuck, Lt. Can feel 'er little pussy twitching. Tryin' tae suck me in."
Another hum. The grip on your asscheek eases as his hand peels away, sliding over swell before notching a finger between your cleft. Dry. Rough. It drags down your seam until it brushes over your fluttering hole, calloused tip digging in.
"soft, too, ain't it?" He asks, words mockingly cruel in their conversational tone. Nonchalant. But Johnny's hands tighten on your waist, palms slick with sweat. Glueing to your flesh. You can tell he likes that. Likes the way Simon talks about you. Demeaning and brutish. Butcher selling a piece of meat. "Bit of a tight fit at first—" he curls his finger inside of you, stretching your sore walls with the width of his knuckle. Sinking in deep. Another follows before you can remember how to breathe around the sting. "But swallows you up like a goddamn dream, Johnny."
His breaths grow ragged. "Fuck, Lt. Look at th'."
It makes you clench up around Simon's fingers, embarrassment scorching through your chest. "Please—"
Neither of them acknowledge you. Simon's fingers split, spreading wide apart as Johnny shuffles forward for a closer look, and nearly choking on his next inhale when he does.
"such a pretty fuckin' pussy—" he says it like a curse. Spitting the words out on a snarl. Angry, now, for reasons you can't discern slobbering over Simon's leg. "God, Lt. ah cannae—"
Johnny shifts back. You hear the clink of a belt. The rip of a zipper. Choked groans barely swallowed down as Simon buries his fingers inside of your weeping cunt over and over again, blunt tips cruelly skating over a spot inside, just behind your navel, that makes you feel liquid and loose between your hips. Debris floating down a whiteriver.
Pleasure peaks with each brutal thrust until you're howling into his leg, unable to move with their hands on your body, holding you down. Making you take it. Making you come undone as Johnny watches.
"fuck, fuck, Lt—she's gonna cum, ain't she?"
"Wanna feel it, Johnny?"
Simon's name falls out of his mouth on a whispered prayer. Drenched in thick reverence. Arched in need.
"aye, sir—" there's something about the hush of his voice, the way it slurs into putty. Enshrining his need in a halo of gold. It sends shivers down your spine. Heats you up fast like a fever. Sends you screaming over the edge—
"gonna miss it, Johnny. She's squeezin' me so fuckin' tight—"
Whatever else they say is swallowed by the keen clawing at the hollow of your throat when you feel the blunt, fat press of his cock knocking against your swollen, stuffed rim.
It's a burning thing—a sharp, heavy ache. Knock, knock. Simon spreads his fingers again, forcing you open. Pulling your hole wide apart for Johnny's engorged head to push up against.
It feels like being split down the middle. Ripped apart. Simon's fingers flex around your nape, thumb brushing soothingly against the knob of your spine.
Can take it, he mutters, brassy and low. A rumble just for you. Gotta take it, birdie.
You forget why. Why you need Johnny's too big, too fat cock inside of your cunt until the head bullies through, scissoring Simon's fingers apart until they're pressed tight on either side of the flared glands. Squeezed between your taut rim and Johnny's cock.
Johnny makes a noise like you've gutted him. A gutwrenching sob. "Oh, shite, Lt. M'—m'nae gonnae last—"
"gonna cum inside 'er, Johnny? Knock my pretty birdie up?"
Right. Right. A baby.
There's a heavy push. Your flesh wrenched apart to fit the fat, throbbing length of his cock—
(the cock that's gonna knock you up—)
Simon's fingers slip out of you as Johnny bucks forward, burying himself deep inside with a long, throaty groan. It's a horrible sensation—a bellyache. Without the splint of Simon's fingers forcing you open wide to near numbness, you're forced to feel the thick girth of his cock. Rim fluttering, spasming over the flared base. Too much, and somehow, not enough.
You sob through it. Each one ripples through your chest until it feels like it will collapse. Every inch of your body burns, throbbing. You don't think you'll survive this ache—
Johnny sets a brutal pace. Likes pistoning into you in quick succession until you're nearly howling into Simon's thigh before slowing to a crawl. Force-feeding you every inch. Making you feel every single one. Long strokes that batter the plug of your womb, bullying against the aching seal of your cervix until the flashes of pain, the savagery of this pleasure, makes you feel sick.
Getting fucked by Johnny like this is both a punishment and a reward. Baptism in hellfire.
Be careful what you wish for—
"gonnae fuck ye 'til it takes, doe. Knock ye up. Want th', don't ye? Aye. Can feel it. Feel this little cunt beggin' fer ma cum. Dinnae worry. Ahm gonnae give it tae ye. A' o' it, doe. Every—fuckin'—drop—"
Each awful word lands like acid on your spine. Chewing through flesh, tissue, until it melts bone below. Liquified. Helpless.
And with Johnny's hands on your hips, anchoring you in place as he hammers into your sore, abused pussy, possessed with the need to carve a space inside of your flesh where only he fits, rots, and Simon's hand on the scruff of your neck, holding you down, there's nowhere to run. Nowhere to escape the ragged breaths that spill from Johnny's slick mouth, the desperate way he pumps into you—thrusts growing sloppy as he stretches towards the precipice they dangle you off of, kicking and screaming as the scent of iron fills your nose, as his flared cockhead scrapes over that place you thought only Simon would ever know. Bluntly battering into the altar that sits, nestled behind your navel, like he's allowed.
Holy offering in a handful of seeds he'll sow over fecund land until something grows.
"Look at you take it," Simon coos, sticky, damp fingers petting over your tear-stained cheeks. It smells of loam. Salt. Iron and ozone. "So pretty when you're gettin' bred, ain't you, birdie?"
It rips a mournful keen from your chest, a feverish moan following on its heels when the lewd squelch, the echoing slapslapslap of Johnny driving into your cunt fills your ears. So wet, so messy, you can feel the slick drying, tacky and thick, on the inner crease of your bent knee.
"He's gonna put our baby in you, ain't he, birdie? Like a good mutt—"
The hands holding you over the precipice let go. Johnny's answering moan spears into your head, fluttering around the pulsing heartbeat of liquid bliss frothing in the pit of your belly. Overflowing over the rim.
Too much, you think, but that's not quite right because you can't feel anything at all except the length of his thick cock lodged deep inside you. Throbbing in tandem with your second pulse.
"gonnae cum, Lt. Gonnae—oh, fuck, Lt—"
His voice is a warm river washing over your spine. Pooling ecstacy. Something heavenly. Divine—
Molten gold blooms in the pit of your belly. Cockhead spitting against the seal of your womb as he cums, filling you to the brim. Fucking it into you even as his cock softens, unable to pull out he says.
Feels like fuckin' heaven, Lt.
"ain't she just?" Simon volleys back, sounding oddly dissonant. Off-key. "Pretty little birdie got what she wanted, huh?"
The drawl of his tone—acid-scorched, electric—forces you to blink through the tears, lifting your aching, wet eyes upwards at him. Searching.
He has the eyes of a predator. Leonine. The gaze of a beast after it's devoured something whole. His touch is as gentle as he can be—a rough, cracked scratch over your blistered cheeks—and when he meets your divining stare, he coos.
"Maybe I'll 'ave a go next time."
In the pounding, soporific slurry of your mind, you can't wrap your head around the words. Can't make sense of them. Struggling to keep your burning eyes open, even.
Not that it matters.
Johnny huffs a scorching breath of laughter over your sweat-slicked spine before wedging his forearm under your belly. Keeping your hips tipped up as he falls into you, resting his broad chest against your back and smothering you into the damp mattress.
"Yer cruel, Lt," he rasps, chin nuzzling over the arch of your shoulder, cock giving a feeble twitch inside of you at something you can't seem to piece together.
"m'jus' givin' my pretty bird exactly what she asked for." Huh? He prods, fingers tapping over your cheek when your swollen eyes slide shut. "Forgettin' y'manners, ain't you? Say thank you, pet."
With Johnny's half-formed chuckle echoing in your head, you mumble the words out on an exhausted sigh.
"an' say thank you to this mutt f'knockin' you up."
It comes out slower this time. Sluggish. His cock gives another twitch as he buries his face between your shoulder blades, smothering a groan.
"Sweetest thing, Lt. Christ—"
"more where that came from, Johnny. Jus' you wait an' see." Another tap. You mewl in response, feeling war-torn and achy. Unable to open your eyes for a second time, all you can do is whimper, burying yourself into his thigh. Pleading, silently, for clemency. Later, you think. Later—
But Simon has other plans.
"Fallin' asleep on me, birdie? Ain't even gonna give me a chance to put my baby in you? Greedy little thing, ain't she?"
Buried under the weight of Johnny as he peppers sucking, open mouth kisses over the width of your shoulder, cum leaking out around the softening plug of his cock, all you can do is snuff out the sob on the arch of his knee, resisting the urge to bite instead.
"Maybe next time then, eh, birdie?" Since you've been so good for this mutt, huh? Maybe I'll give you a reward.
Just be careful what you wish for, huh, birdie.
#i don't know how to end things sorry#simon riley x reader#johnny mactavish x reader#soap x reader#ghost x reader
3K notes
·
View notes
Text
I know we always talk about Garroth ending up looking exactly like his father, but what about Dante growing up to look eerily like Gene.
When he joins up with Phoenix Drop, he's still young. He's a little on the short side, still a bit too thin from life in the wild and imprisonment, and he's a little anxious and shaky around so many people after having grown unused to living in a village. The smiling faces of the citizens remind you of your old home, of clamoring crowds and standing frozen in the plaza as your brother . . .
Anyway, it's good here. It's easy to fit in. The guards joke around with you and make sure you're healthy. They don't know a thing about dual wielding, but you get plenty of sparring partners out of helping the local baker practice her magick, and you maybe make a friend too. You're not too sure how you feel about the Lord, but she's a kind soul and does her best to make sure you're comfortable here in town, and her kids are great. Babysitting the boys is easily your favorite duty. Yeah, it's good here. For the first time in a long while, you feel like you're doing good.
Then the war comes. The children and non-combatants are sent away. The jovial atmosphere of the guard tower has soured into solemn silence as you make your final preparations. In the morning, you step into the battlefield and you go to war for the first time in your life. You have a horrible feeling in your gut that it won’t be the last.
You, Sir Laurance and Sir Garroth make a good team. It makes you sick. The three of you cross the battlefield at a slow and inevitable pace, cutting down any soldier that dares stray too close, and together you cleave the enemy forces in half, scattering them. The killing comes easy to you. You had hoped that in this peaceful new village, with time, you would become unfamiliar to how easily you were once able to take a life, but right then you’re glad your body never forgot the motions of death. Glad for the blood that stains your hands—how can you be glad?
You can’t remember how long you fought for. Days, weeks? Surely not months, or so you think. Yours is a small force, and though Miss Lucinda is a good healer, she grows tired while the other army’s numbers are replenished time and again. You remember the bags under her eyes as she tipped a potion sip by sip into your mouth the time you were shot through the face.
You remember sneaking into the enemy camp in the dead of night, skirting around the edges of it to the back line where the archers rested. You quietly slit five of their throats before you were noticed, and managed to slash another across the belly before the arrow caught you in the side of the face, in one cheek and out the other. The wood of the shaft cracked when you bit down. It was everything you could do not to scream as you fled. Dale thought you were a fiend when you first stepped out of the shadows, face obscured in blood and cradling your jaw as you cupped a hand beneath your mouth in an effort to catch more blood before it left a trail. Laurance held you while Garroth split the arrowhead from the rest of it with a knife and pulled the shaft out the other side of your face, your jaw gripped tight in one hand to keep you from struggling. It took hours to pull the splinters from your cheeks and tongue before they sent you to wake the healer. The whole ordeal had been excruciating. You might have cried. You remember that a lot more clearly than most other times at war. After a while, it’s hard to tell which side spills more blood when so much is shed that red squishes out of the earth wherever you step.
Every day, you fought dawn to dusk. And then one day you won. By Nicole literally knocking some sense into her father, of all things! You find a quiet corner to throw up in and for a beautiful moment, you think life in this little town you’ve started thinking of as home will go back to being good. Until your Lord tells you to guard the village as she races past the gates, and she doesn’t come back. None who followed her do either.
For days, you stand waiting at the gates. You don’t eat, you don’t sleep. O’khasis is gone, Scaleswind has made a refuge of the plaza, and still there is no sign of your Lord or your brothers-in-arms. You won’t even leave to have your wounds seen to. Nicole has to drag a doctor to the gates to treat you, and the entire time you watch the forest hoping that any moment they will reappear. You only step away when someone brings you news that the ship that took the children away has returned. You should be the one to tell them.
Zoey knows something is wrong the moment she sees you. Levin and Malachi smile and ask where their mother is—they call you ‘uncle’ while they do. You get down on your knees before them, and you gather them close in your arms, and you cry as you tell them their mother has been missing since the day the war ended. You’re still holding them when the exhaustion catches up with you.
Zoey is with you when you wake. She tells you you’ve been out nearly two days. She fusses over you, and you know you’ve worried her because that’s what she does when she’s worried. You’re a mess anyway, so you let her fuss. You drink the broth she makes you, you change into the clothes she provides, you sit still while she cuts the unruly mats of your hair and shaves your face. You used to cut yourself shaving all the time, no one ever taught you how and you were only six or so when Gene was learning to; you don’t remember now how he showed you each step or the laugh in his voice at the face of disgust you made when you slapped a little hand into the lather on his face and left behind a tiny palmprint. Zoey doesn’t cut you once. When she’s done with you, she takes you by the arm and guides you back into civilization, where everyone who remained has decided already on search parties to go out looking for your missing friends.
You head each expedition. Dale brings himself out of retirement to watch over the town while you’re gone, and asks only that you also look for his son. Does he know you used to be a tracker, used to spend days in the woods trailing coyotes and runaways for enough coin to carry you through the cold months? You try for him, but the ground is soft still and every step anyone takes leaves a print, all overlapping and muddled. You keep an eye out as you circle the same stretches of woods for days, but you find nothing. Your group goes further and faster than any other, the first to find and dismantle bandit camps and dens of fiends, but no matter how far you go you find not a sign of anyone who has disappeared that day. It’s as though they vanished into thin air. Every time you return home, Dale looks at you with hopeful eyes, and every time you must take him aside and break his heart a little more. Eventually, he stops asking.
For a year, you search. The area has never been safer. You have never felt so alone as when people start to suggest that a funeral may be in order.
You feel like a monster for the rage in your voice when you denounce these people. You know they aren’t dead—you would have felt such a thing, you know, you would have felt pieces of yourself snapping like wire pulled too taut, you would have felt the sharp edges tangling inside you—it would have felt like it did when the brother you killed rose from the grave to slit your throat and cut your very existence from the memory of Boboros. You hear white noise rumbling in your ears when the first brave soul says Sir Dante, there’s been no sign for a year now, and your blood is boiling when you slap their comforting hand off your shoulder. You spit that you’re not giving up just because everyone else has taken no evidence of life to mean the surety of death, and with their pitying looks burning into your back to return to the woods. You scream into the trees until you can’t anymore. When it doesn’t help, you use your considerable tracking skills to hunt something, anything, until you feel human again.
You crawl back home the day before the funeral with your cape stained with blood; they held it back so you could attend. You polish your armor and swords until they shine, and the warped reflection of your own face makes you feel sick the way waging war did. You stand at attention the entire ceremony without moving a muscle. When Dale reads the names of the deceased at the end, offering their souls into the embrace of the Matron, you salute, and the clatter of your armor silences the crowd.
Everyone who fought in the war salutes with you. So do your Lord’s sons. You’re too tired to cry. You hold your salute long after everyone else has left.
The remaining forces of Scaleswind return home. One by one, family by family, the streets of your home empty. Without your Lord, without your guard, the citizens trickle out the front gates and never turn back. Some apologize to you as they say their goodbyes, and some of them you actually believe. You close the gate behind each of them until all that remains is you, Zoey, and your Lord’s sons. Then Zoey tells you she’s taking the boys to the Yggdrasil Forest. She holds you tight for too long and kisses your brow when you show them to the gate for the last time.
You can’t believe you ever thought you knew what loneliness was before this.
For five years, you are completely and utterly alone. You search and you patrol and you do your best to maintain the village. You don’t believe in Irene, but every day before dawn you stand before her statue and look down down down over the cliff’s edge and pray that this won’t be the rest of your life. That you haven’t deluded yourself into believing a fantasy, that you haven’t made such an incredible fool of yourself that people can’t bear to be around you, that you haven’t been forgotten. For five years, you pray that someone, somewhere, remembers that you exist. You look down down down over the cliff’s edge and have the terrible thought that you don’t know what you’d do if you were forgotten again.
The gate is falling apart. You don’t know how to repair the damage the weather’s done to it, you tried to patch the cracks but it never holds. With each year, you’ve been pushed further and further outtowards the coast. The only places you have the energy to maintain anymore are the guard tower and your Lord’s home. You blockaded the gates when the mechanism broke, you check it on occasion to be sure no bandits get in, and one day you hear voices from the other side. Familiar voices. You scramble up the wall and look over the other side at a boy you don’t recognize looking back up at you. He says, Is that Uncle Dante? and you climb down as fast as you can to embrace Malachi.
He’s nearly the age you were when you first met his mother. He’s grown tall, and strong enough to carry his brother on his back. Levin is fevered when you first see him, flush and hurting even as he dozes, and Malachi tells you he can’t walk from how bad he hurts. You remember how Zoey fretted over him when he was young, how sometimes he’d scream for seemingly no reason, and once you show them to their mother’s home Malachi refuses to leave his bedside.
You sit with them and ask where Zoey is. Malachi tells you of her obsession, and the relief that you are not alone in the belief that those who disappeared are alive feels like a hint of betrayal. You’re relieved that she’s driving herself into a downward spiral because of what? Because it makes you feel like you were reasonable to fight not to let their souls be put to rest?
You wait for her at the gates deep into the night and take her to her boys when she bursts from the woods, frantic that she’d lost them, and safe if your Lord’s home she holds you so tight your ribs hurt from the force of her grip. After so long, you’re not alone anymore.
You wake before dawn and strap your swords to your back. For the first time in a long time, you feel safe enough to go without your armor. You hike up the steep cliff to the Irene statue. You kneel before her to offer your thanks. You look into the pool at her feet and fear grips you by the throat.
Your brother’s face looks back at you.
You wear your swords the way he did. Your hair falls like his, dark in the shadow of Irene. Your face is gaunt and pale from old habits, eating only enough to sustain yourself so rations will stretch long enough for you to find more—do you remember how they starved Gene before they killed him? How they weakened him so he wouldn’t have the energy to fight? How pale and gaunt he was, dirt streaking over the side of his face, blood and grime drying in his hair, shaking and sweaty with how hard he fought back? Do you remember the scar that twisted around his throat when he returned from the dead to get his vengeance? Your collar is open over the scar he left twisting across your own, and it matches his own so very well. In the shadows of your eyes, you see his own staring back.
You think of the war. You think of how easy the killing was. You think of how easily Gene cut through the guards, the Lord, the memories of Boboros. The rage in his voice when he denounced you as his brother, the twist of his smile when he told you he would leave you to rot, Dante. No one will ever remember you. You can see that twist in the corners of your own smile, pushed into shape by the deep scars on your cheeks. You and your brother are the same.
You’re shaking too much to stand. You never go without your armor again.
#do you think growing a beard helps any#aphmau dante#mcd dante#dante the forgotten#dropofsunlightextras#loyalty of memory#mcd#aphblr#aphverse#aphmau mcd#mcd rewrite#minecraft diaries#aphmau minecraft diaries#mcd gene#oof long post#mcd malachi#mcd levin#mcd zoey#zoey taltatheil#mcd dale#tw blood#tw abandonment#tw injury#tw violence#tw implied death#tw war mention#I let him think for two seconds that everything's good and will be back to normal any day now and then I punch him in the face#kuri writes#I love Dante and the potential for making him break himself down into his fundamental pieces only to find that they match Gene's#honorable mention of my disabled Levin headcanon
143 notes
·
View notes
Text

Extracts from Gio’s diary.
a little transcript in case my handwriting is too terrible:
august 2021 (before first breakup with aaron)
you are expanding and constricting in blues and purple, bruised lungs, beaten and deoxygenated above me. me, trapped in your ribcage. i feel like the body who’s spleen i sleep on had swallowed a stone, and i’m just rattling around your torso.
i wish i was buried between someones vertebrate but theres no one around to eat me. i dont think im a very a pretty stone, maybe id be more edible if i were smaller or if i were a kinder stone. i dont think people would care that i was hard to swallow then. But im not a pretty stone and im not a smart stone so they feel every scratch as i drag myself down their oesophagus.
I think it would be better if people got operations and surgeries to remove me anyway. i dont want to be in there anymore, i want to turn faster and farther in between their organs so they take me out. i dont really want them anyway. i want to be a stone on the shore, i dont think id be on a shingle beach. i dont know if there are any stones anymore, i think im the last to be removed. ive been feeling sick a lot more recently, my heart beats so fast and heavy in my chest. it shocks me
i think sometimes its easy to forget im alive, im just going through the motions
october 2021 (just moved schools)
“I am not an earth nor an adjunct of an earth / I am the mate and companion of people, all just as immortal and fathomless as myself”
I cant begin to understand him, his heart is pouring out love for everyone that passes him, how could he feel more connected to uncaring strangers than uncaring nature? the stranger chooses not to care, chooses to loathe and anger, nature has no say in it. It creates storms and tsumanis and fells trees and cleaves land in on itself through no fault of it own, through no choice. Maybe the stranger doesnt have a choice either, maybe that is nature, a passage of self and outward destruction writen into genes and dna and into lines of bark and blades of grass. Maybe its nature not to care, but that makes me too sad to believe. and anyway, if i was to be hurled into an uncaring sea or an uncaring crowd of strangers, id ask to be ploughed against rocks and turn sea foam pink, i dont like strangers.
I need to stop thinking about that stupid passage about his stupid abundance of understanding. I need to read something else, probably do some homework or something.
#my ocs#oc#ocs#original character#writing#original characters#oc artwork#oc art#original charater art#art#ignore the fact some of this is taken word for word from my old diary#we dont need to talk about that
13 notes
·
View notes
Text
Where's Everett? (1966) Or: Alan Alda raises an invisible alien baby
Part of a series of writeups of unaired or otherwise inaccessible TV objects ft. Alan Alda, which I was able to access via the Paley Center for Media in NYC (November 2024).
Where’s Everett? (1966) - Producer, Creator, Writer: Ed Simmons; Associate Producer: Jon Zimmer; Director: Gene Nelson.
Cast: Arnold Barker - Alan Alda; Sylvia Barker - Patricia Smith; Dr. Paul Jellicoe - Nicolas Coster, Lizzie Barker - Doreen Miller; Murdock - Frank DeVol; Milkman - Robert Cleaves.
Summary: When a basket containing an invisible baby appears on the doorstep of Arnold (Alan Alda) and Sylvia Barker (Patricia Smith), the couple clashes over what to do. Arnold, a professional science fiction author, thinks they ought to turn the baby over to be studied. Sylvia thinks they ought to raise it as their own. The couple call in family friend and pediatrician Dr. Paul Jellicoe (Nicolas Coster) for help, all while trying to balance keeping invisible baby Everett a secret from their daughter, Lizzie (Doreen Miller), nosy neighbors (Milkman - Robert Cleaves) and investigating reporters (Murdock - Frank DeVol).
Plot recounting: The show begins in an unassuming suburban American neighborhood, white-picket-fence’d, green lawn’d, and quiet. A tracking shot follows an empty, floating wicker basket down the street, through a white picket fence, and up to the door of a house. The basket floats down to sit on the doorstep. The doorbell is rung by an unseen entity, whose footsteps are audible as it hurries away. The camera follows the invisible entity back down the street, panning to reveal a large spaceship parked on one of the suburban lawns. The spaceship opens, a staircase descends, footsteps are heard ascending it, and the staircase retracts. The spaceship flies away. Meanwhile, the doorbell is answered by Arnold Barker (Alan Alda), still in his dressing gown. He regards the empty basket with confusion as a baby—not visible—begins to cry. The intro plays: A cheerful, upbeat tune over yellow title cards listing cast and credits.
Arnold and his wife Sylvia (Patricia Smith) investigate, attempting to locate the crying baby. They decide the sound must be coming from inside the apparently empty basket. Though Arnold is doubtful, he puts a hand inside the basket, and quickly withdraws it: He was bitten by something invisible, which left visible teeth-marks (“Arnold, there’s a baby in that basket!” “But a baby what?”). The couple try to decide what to do with the invisible baby. Arnold suggests they report it (“Why don’t we call the police and have them give the baby back to Claude Rains?”), but they eventually settle on calling their friend, pediatrician Dr. Paul Jellicoe (Nicolas Coster) for advice.
Arnold explains the situation to Paul, who refuses to believe him, due to the fact that Arnold is a professional science fiction novelist. Eventually Paul agrees to come over for Arnold’s sake, convinced that Arnold is sick, drunk, or otherwise hallucinating. Meanwhile Sylvia feeds the baby, whom she has named Everett, oatmeal, which disappears off the spoon. Arnold complains about the ridiculousness of the situation, and Everett responds by flinging oatmeal into his face.
When Paul arrives, Arnold shows him to Everett’s wicker basket—but they realize that Everett has crawled away, prompting a carefully thorough and shoeless (“You might step on him!”) search of the floor and furniture, until Paul notices a floating piece of candy and realizes Everett the invisible baby is real after all. Paul gives the baby an exam and notes that the child is healthy and normal, except for an unusually high body temperature and, of course, the invisibility (“Just think, Arnold, you’re going to save a fortune on clothes.”). He recommends the Barkers not mention the baby to their young daughter Lizzie, worrying it might have an adverse emotional impact.
Lizzie descends the stairs and states that she dreamed she heard a baby crying. The Barkers and Paul play a game of hot potato with invisible baby Everett, trying to keep him away from Lizzie without arousing her suspicion or looking too obviously as though they are holding something invisible and baby-sized. The tradeoff only works until Everett is placed back into his basket and begins to cry. To the surprise of the Barkers and Paul, Lizzie takes the revelation with equanimity: “Invisible baby,” she says, pleased, before rocking Everett in her arms and asking if the family can keep him.
Sylvia hears the milkman outside, and asks Arnold to buy some extra milk for the baby. Arnold, still believing they ought to turn the baby over to somebody in authority, reluctantly agrees, and hears from the milkman that a rumor is going around the street: A flying saucer was spotted on a neighboring lawn early in the morning.
Arnold is surprised and then delighted by the news, and suggests that it could be a boon for his career. He proposes handing the baby over to science in a public fashion, imagining the headline attention it might receive, and the increase of attention towards his books. Sylvia, however, wants to keep Everett, distressed by the idea of the baby being “poked at” and studied; she states that whoever dropped the baby off must have wanted them to care for him.
The doorbell rings (Arnold: “If nobody’s there, maybe it’s the parent.”), and reporter Murdock (Frank DeVol) asks if the Barkers have any information about the flying saucer rumor. Arnold attempts to tell the reporter about Everett, but Sylvia and Lizzie undermine him by implying he is relaying a story he’s working on, mentioning his career as a science-fiction writer, and hiding the baby, so when Arnold directs the reporter to pick Everett up, he finds nothing in the basket. Arnold immediately panics, believing Everett has crawled away again (“Where’s Everett?!”), and begins to search the floor on hands and knees. The reporter leaves, convinced Arnold is insane.
Sylvia and Lizzie assure Arnold that the baby isn’t lost again—they only relocated Everett to an armchair. Arnold picks Everett up for safekeeping and tries to convince his family one last time that they can’t possibly raise an invisible baby. He slowly begins to doubt himself as he speaks, noting that they have always wanted a son, before Everett manages his first word: “Dada.” Arnold leaps over a couch and begins to hop around, delighted, holding Everett to his chest (“He knows me!”), accepting his role as Everett’s new father.
(Semi-Alda-specific) review: So awesome. Went into this one knowing the premise “Suburban couple find and raise invisible alien baby” and was prepared for it to be pretty hard to take, but was pleasantly surprised by its consistent humor and how well it committed to a pretty ridiculous conceit. Ed Simmons wrote for Martin and Lewis, which feels just about right, given the zaniness of the script and the prevalence of tossed-off one-liners. Unclear on how this would have worked for longer than one episode, but I’m sad we never got to see it attempted. Alda and Smith are particularly excellent at what seems, while watching, like a long-form improv warmup game: They commit and commit well to ‘holding’ baby Everett, transferring him between each other, rocking, soothing, lifting, and otherwise gently handling baby-shaped empty air, which if you stop buying the existence of the baby becomes insanely funny insanely fast, but in the watching process is pretty sweet. Really excellent for Alda specifically because he does befuddled but well-meaning and generally reasonable suburban dad so well as a role type; plus he’s wearing a wonderfully 60s velour tracksuit top, which is so visually fun it’s almost a character in itself. Verdict: Don’t miss this one if you visit the archive. Such a treat.
#alan alda#where's everett? (1966)#lost media#<- not really because archived but etc etc#paley center#ed simmons#1960s TV
10 notes
·
View notes
Text
SARS-CoV-2 can infect and replicate in human motor neurons differentiated from induced pluripotent stem cells - Published Jan 4, 2024
Numerous patients experience neurological and neuromuscular symptoms during and after infection with the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Apart from the central nervous system (CNS), the peripheral nervous system (PNS) is also affected. In this study, the Italian authors investigated whether SARS-CoV-2 can infect and replicate in human motor neurons (MNs) differentiated in vitro from induced pluripotent stem cells (iPSC-MNs). They also examined whether iPSC-MNs express the main receptors for SARS-CoV-2 entry and whether SARS-CoV-2 infection of iPSC-MNs changes the expression of 46 genes involved in cell survival, metabolism, inflammatory response, apoptotic and antiviral pathways.
SARS-CoV-2 is an enveloped, positive-sense, single-stranded RNA virus. Its genome encodes four structural proteins, namely the spike (S), envelope (E), nucleocapsid (N), and membrane (M) proteins. It seems that SARS-CoV-2 uses various neuroinvasive strategies and entry pathways to invade the nervous system, such as infection of the nasal olfactory epithelium and axonal transport along the olfactory nerve, retrograde axonal transport, invasion by compromising the blood-brain barrier, retrograde virus spread from the lungs to the CNS via the vagus nerve, and the use of infected hematopoietic cells as “Trojan horses” (hematogenous route).
It remains unknown whether observed neuromuscular manifestations of SARS-CoV-2 infection are caused by a direct viral invasion of motor neurons, and/or are a collateral injury resulting from an uncontrolled innate immune response. Damage to motor neurons leads to the deterioration of muscle function, manifested as muscle weakness, atrophy, or paralysis.
Two host-cell factors are important for SARS-CoV-2 viral entry into many cell types: angiotensin-converting enzyme 2 (ACE2), which is bound by the S protein, and transmembrane serine protease 2 (TMPRSS2), which cleaves the S protein, allowing this binding to take place. In addition to ACE2 and TMPRSS2, the S protein has been reported to engage other cell-surface factors proposed to serve as attachment factors promoting SARS-CoV-2 entry.
The expression level of ACE2 is low in the human brain. In contrast, neuropilin 1 (NRP1), CD147, TMPRSS2, and furin are higher and broader expressed than ACE2, indicating that they may be putative mediators of SARS-CoV-2 entry into human nervous system cells. According to the authors of this study, the infection of human motor neurons with SARS-CoV-2 mainly relies on CD147 and/or NRP1 binding. Previous studies have shown that NRP1, known to bind furin-cleaved substrates, potentiates SARS-CoV-2 infectivity and that the furin-cleaved S1 subunit of the S protein binds directly to cell surface NRP1. www.science.org/doi/10.1126/science.abd2985
About the study The authors used an in vitro model of human motoneurons (MNs) differentiated from induced pluripotent stem cells (iPSC-MNs) to investigate the infectability of these cells by SARS-CoV-2 and their expression of the main SARS-CoV-2 receptors. They also examined whether SARS-CoV-2 changes the expression of 46 genes involved in cell survival, metabolism, inflammatory response, and apoptotic and antiviral pathways.
The induced pluripotent stem cells (iPSCs) from three healthy donors (1 male and 2 females, aged between 37 and 49 years) differentiated into motor neurons that expressed both, neuronal (bIII-tubulin and SMI-312) and motoneuronal (ChAT, HB9) markers.
To verify the infectability of iPSC-MNs by SARS-CoV-2, VeroE6 cells were exposed to supernatants collected from iPSC-MNs infected with SARS-CoV-2. Reverse transcription polymerase chain reaction (rt-PCR) was used to analyze the expression of SARS-CoV-2-specific ORF7A, ORF3A, ORF8, RDRP, S, E, and N genes in uninfected (mock) and infected iPSC-MNs.
The expression of main SARS-CoV-2 receptors (ACE2, CD147, NRP1) and peptidases (TMPRSS2, furin) was assessed in iPSC-MNs infected with SARS-CoV-2 and the A549-hACE2 cells, as a positive control.
Results To validate the infectability of iPSC-MNs, VeroE6 cells were exposed to supernatants collected from infected iPSC-MNs at different time points. The results showed that supernatants collected from infected iPSC-MNs could re-infect VeroE6 cells. Furthermore, the SARS-CoV-2-specific genes (N, E, S, ORF3A, ORF8, and ORF7A) were detected exclusively in infected iPSC-MNs. The expression of SARS-CoV-2 N1, S1, S2, and E2 genes in infected iPSC-MNs was subsequently confirmed by rt-PCR.
The expression of SARS-CoV-2 specific N1 and N2 genes was identified in supernatants from infected iPSC-MN cultures by rt-PCR, confirming that SARS-CoV-2 can infect and replicate in human motor neurons. However, the viral replication level was lower in iPSC-MNs than in VeroE6 cells. In addition, SARS-CoV-2 replication in infected iPSC-MNs was not accompanied by a cytopathic effect as assessed by the crystal violet assay. In addition, the immunofluorescence assay detected N protein in infected iPSC-MNs, mainly at the perinuclear level, in the soma, and along the neurite extensions. However, the percentage of infected iPSC-MNs was very low.
Further analysis demonstrated that iPSC-MNs expressed the main entry receptors of SARS-CoV-2, including ACE2, CD147, NRP1, and TMPRSS2, but at different levels. ACE2 and furin were expressed at lower levels, whereas CD147 and TMPRSS2 were expressed at higher levels in infected iPSC-MNs compared to the control A549-hACE2 cell line. The NRP1 expression was comparable between iPSC-MNs and A549-hACE2 cells. The immunofluorescence assay for ACE2, CD147, and NRP1 confirmed these results.
In iPSC-MNs, SARS-CoV-2 infection changed the expression of 10 genes involved in cell survival, metabolism, antiviral, and inflammatory response. The virus up-regulated the expression of B-cell lymphoma-2 family protein (BCL2), BCL2-associated X protein (BAX), caspase 8, CD147, proinflammatory interleukin-6, and sphingosine-1-phosphate receptor 1, involved in the regulation of lymphocyte trafficking, brain and cardiac function, vascular permeability, and vascular and bronchial tone. The virus down-regulated the expression of human leukocyte antigen-A and endoplasmic reticulum aminopeptidase 1, involved in antigen processing and presentation, and angiogenin, which exerts neuroprotective functions and contributes to the systemic response to infection.
Interestingly, an increased ratio between the expression of anti-apoptotic BCL2 and pro-apoptotic BAX gene suggests that programmed cell death was somehow prevented in iPSC-MNs after the infection.
Conclusion The authors concluded that this study has shown, for the first time, that SARS-CoV-2 can infect and replicate in iPSC-derived human motor neurons. However, viral replication and the percentage of infected cells were lower than in VeroE6 cells, susceptible to SARS-CoV-2.
This article was published in Frontiers in Cellular Neuroscience.
Journal Reference Cappelletti G, Colombrita C, Limanaqi1 et al. Human motor neurons derived from induced pluripotent stem cells are susceptible to SARS-CoV-2 infection. Front. Cell. Neurosci, Sec. Cellular Neuropathology. 05 December 2023. Volume 17, 2023. (Open Access) doi.org/10.3389/fncel.2023.1285836 www.frontiersin.org/journals/cellular-neuroscience/articles/10.3389/fncel.2023.1285836/full
#mask up#public health#wear a mask#pandemic#covid#wear a respirator#covid 19#still coviding#coronavirus#sars cov 2
5 notes
·
View notes
Text
Ok but what if bc Lucien has strong magic genes from his mother and the spell-cleaving genes from his father plus that all-seeing eye he can break mating bonds and he breaks the feysand mating bond and sparks chaos. Should I write feral Lucien?😈
#pro lucien vanserra#lucien vanserra#my ideas are getting more and more evil#dare i write it#lucien spell cleaver#anti feysand
43 notes
·
View notes
Note
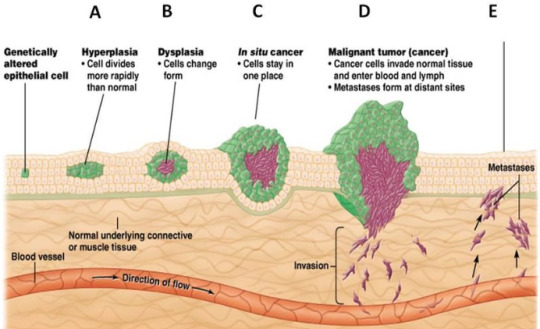
Random ask, can I ask what are apoptosis and caspases (in simple terms)?
I have been summoned
(my 'noorie rambles. be very afraid' tag will make so much sense now. I tried to talk about it in a way that's a way that's both engaging and simple. Lots of metaphors.)
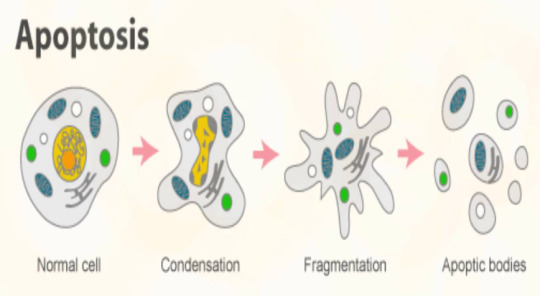
Caspases destroy the cell if it needs to die. They dismantle the cell parts as the cell membrane turns into bubbles, containing all the damaged pieces within so that a white blood cell might disposes of them. That's apoptosis in a nutshell.
But why must the cell die you ask?
Plenty of reasons. Human embryos have webbed fingers, and apoptosis causes the webbing to dissipate. Tadpoles too, use apoptosis to rid of their tails so they can become frogs. Apoptosis opposes growth - aka mitosis - and it keeps the body in homeostasis - or a balance of sorts. The destruction to mitosis's creation.

However, what I find most interesting, what is the subject of much research nowadays, and what holds the most opportunity for angsty biology fanfics (most important/j) is that apoptosis opposes tumorigenesis. In other words, if something goes wrong, if the cell gets some notion that it might try to cheat death and live forever, if it has the inklings of an idea to hijack the rest of the body through growing its own lump of cells and draining the body of its nutrients... if it decides to throw a mutiny at the expense of peace...
Then the cellular system realizes, and it activates the caspases.
(Usually how a cell 'decides' is when something wrong happens with genetic replication, mutations and such and such. There are two labels of genes relating to this. Proto-oncogenes promote cell growth and avoid apoptosis - these are the creation genes, the ones that wish to achieve the heights of production and throw all caution to the wind. Whereas tumor suppressor genes are... tumor suppressors. They are the little 'angel on the cell's shoulder' that says 'you've done wrong, now commit cell death and accept your fate'. When a proto-oncogene mutates, it becomes an oncogene. Basically, the genes that said to grow for the sake of the body now says to grow at the expense of the body, the 'devil on the cell's shoulder' won... and now it'll try to cheat death)
Caspases are a type of enzyme, which is a subset of proteins. I like thinking about it like this: enzymes are witches, spell crafters. Proteins in general function like worker bees, but enzymes are the ones who actively create and destroy, the ones that change the way the universe manifests so life can exist. They utilize reactants from their environment - materials which they are named after, such as proteases that cleave other proteins - to either create larger, energy storing molecules (endergonic reactions), or break down large, energy storing molecules to release energy (exergonic reactions).
(One type of enzyme that I like is called kinases, they basically initiate things, they tell the cell 'it's okay to do things' or 'this will be dangerous, let's not do that', or 'oof this cell doesn't have the proper genes, that could be a problem, how about we destroy everything so this doesn't become a big issue?')
When the cell needs to die, the caspases activate.

Humans have 14 caspases. I wish I knew what all of them are, but the pub med articles I read do not have a whole list of them like a pokedex from pokemon (at least that I have read so far, after ap testing there's one article I want to read that seems to have ANSWERS to a lot of my questions)
So imagine this, the cell has lots of caspases floating around in the cytoplasm/water jelly environment. These beings of death in huge numbers in a perpetual slumber, just floating around, until the cell decides it needs them to destroy its existence, and it activates them.
(ALTHOUGH some of my questions refer to the "caspase-dependent non-lethal cellular processes", so far what I've been able to find is that if the cell needs some repair to the cytoskeleton/structure of the cell, then it activates a few caspases to destroy parts of it, so that the repair may start.)
There are two types: inflammatory caspases (if I remember correctly, this is caspase 1,4, and... a few others I'll have to check) and apoptotic caspases. What we're interested in are the apoptotic caspases. Initiator caspases get activated first. The way I story-fy them/see them in my head is as the older sibling types who condone violence. If the cell needs to die because of an external signal - a message from far away to destroy itself, to which it must oblige - then caspase 8 activates. If the cell needs to die because of an internal signal - something inside isn't right, and what a havoc it would be to pass that on through replication, so the proteins decide the fate of the world it lives in, and it choses death for the sake of the wider body- then caspase 9 activates.
Both these initiator caspases cleave (meaning 'cut', these inactive beasts are not yet 'complete', so they need further modification after activation to work properly. If the cell were to create caspases fully functional and finished, the large number of caspases would kill the entire cell, so they don't finish, they subdue the ones behind their death, and make sure to regulate their slumber.) their 'younger, violent siblings': caspase 7 and caspase 3. The doll I have is a caspase 3.
(I've found more information on them than caspase 7... although.. the one article I found.... it should have information on them all, the amount of searching I've done for those sorts of answers... but alas... I need to read the material for the ap bio exam... which does not include caspases.... one day...)
Caspase 3, as far as I could gather, destroys the cytoskeleton. The cytoskeleton supports the entire cell, acting as the 'tent poles' that keep the floppy cell membrane from collapsing. Collapsing, however, is exactly what the caspase 3 wants. It dismantles the whole thing, along with activating and inactivating a slew of other proteins. (it's an assassin basically.)
And with this, the caspases bow, for the cell is separated within these small little 'blebs' (they look like bubbles) and the cell is destroyed.
OH AND LAST LITTLE THING!
If something inhibits the caspases, the cell goes through with necrosis, which is basically instead of becoming little bubbles, the cell membrane ruptures and the cell 'guts spill out'
THANK YOU SO MUCH FOR THE ASK!! <3 <3 I got to blabber about caspases and it gave me happy sparkles.
ALSO, here's the doll I made for caspase 3:

#biology#scientists#science#apoptosis#enzymes#caspases#science side of tumblr#science side of the internet#noorie infodumps. be very afraid#noorie answers asks#lobotomy for my brainrot
9 notes
·
View notes
Text
by happenstance I read this poem today and now it's rattling around inside my skull like it has the place to itself
(if you're a cryer like me, be warned)
ALZ Ghazal
By Rebecca Foust
For my sister
It’s the same house, same rugs, same wallpaper, and bedroom repeating;
same dresser; same rocker. Same window and frame, repeating.
Same birds at the pane, same pots and pans, and—on the alarm clock,
the wall clock, the phone clock—the same time, repeating
each hour’s increment in a lived life. But, This is no life, each day like
before and to come, repeating.
The furniture set in a known pattern. The rugs there, like always, inking
the blueprint of home, repeating
jewel tones on the floor, but what was once north–south now seems to lie
east–west—who moved the rugs?—in sum, repeating
the familiar, but sideways. Your inner axis has shifted, the landmarks
somehow changed but the same, you repeating
Why do they keep moving the rugs? The desk, the chair, your keys?
Home its own balm, repeating
the familiar, but neither keys nor your purse can be found—I know
I just had them—repeating
the questions yields the same, that is, no real answers. Your sense of taste
gone, like eating chum, repeating
the same million small motions: fork to plate then mouth, then back down,
always the same, repeating
the flavor of cardboard. You used to love to cook, that joyous jazz variation-
on-a-theme now a repeating
like pages of musical staffs, xeroxed blank with no notes. Lately, you refuse
to eat anything at all. In a poem, repeating
lines compose a refrain and, echoed again and again, the sum of refrains
is a song. But there is also empty repeating:
zero plus zero plus zero still zero, a void. No accretion, no growth, no life,
no thrum. Then again, birds—some, repeating
one clear note, are said to singing without tune—and, the same set of sounds
from a beaten drum, repeating
means nothing and everything at the same time. The gene runs in families
and can be followed like breadcrumbs, repeating
the precise map for getting lost, down through generations. She took the same
route to work on the town tram, a repeating
my sister relied on. We rely on a plum to taste purple when our teeth break
its skin. Some numbers go on ad infinitum, repeating
that reminds us of architecture and God. Rings of a tree, whorls of a shell,
sections of a lime, repeating
a sequence that re-enacts growth and change. Meiosis, mitosis, Eve cleaved
from Adam, other kinds of repeating
so life can go on. We are “pattern-seeking animals,” Hass says, and even
atheists pray in a jam, repeating
lessons learned as a child: “Now I lay me” like the promise of mercy, like mercy
itself, soothing and warm, you repeating
your talisman against terror. After your nap, I worry you’ll wake confused,
but then you resume, repeating
Where are my keys? Why do they keep moving the rugs? Always theft and loss
the theme, repeating.
They say you’ll forget my name, and your own. How to talk, small mercy
then, you no longer doomed to repeating
your aching questions. They say you’ll forget how to swallow, to breathe—
O God, will you forget to breathe?—just succumb, repeating
the pause in place of the heartbeat? The doctor says keep things simple
and flat, no drang or sturm, repeating
“Is it nice there today,” and “Have the tulips bloomed,” conversations dull
with the same datum repeating
catechism and cliché. You no longer work on your dollhouses, potting trees
the size of your thumb, repeating
postage-stamp paintings on walls, building one-inch-to-the-foot the lavish life
you never had or presumed, repeating
a litany of loss enlarged from what-was to what-will-never-be. The Christmas
tree shadowbox in simulacrum repeating
the childhood obsessions. Some of your dollhouses have tiny dollhouses
within them, repeating
themselves in infinite regression. All that helpless sap and surge of creation,
your xylem and phloem repeating
what to make of a diminished thing. You say you’ll burn them come spring,
without reflection or qualm, the future repeating
and eating the past. It’s happening so fast: a page torn, a book lost, then
whole libraries gone dumb, repeating
erasure until only erasure remains. O trochee, lilting your 10-beat line.
O river, O iamb repeating
the tide. Bishop called loss an “art.” I last saw you at home in October,
a glorious autumn repeating
saffron and scarlet against azure sky. Outside now, a robin taps at the pane,
ravished by her own image, an eye-rhyme repeating—
there’s been one bird each spring. Some beat their beaks bloody and die,
some stay for days, numb with repeating
their broken routine. Who moved the keys? Who moved the goddamn keys?
Your patient husband is grim, repeating
they’re-in-a-safe-place-so-we-can-keep-you-safe. We are all of us trapped
under a great dome repeating
our words and the cries of wild birds. This is harder in some ways, he says,
than Vietnam, repeating
his pledge to stay till the end. He tries to give you small pleasures, picking
and handing you the plum repeating
all you can recall of lush purple juice run down your chin, but at best,
it’s a dim meme repeating
as holograph plums eaten before. Soon, where are my keys will dwindle
to where keys, the totem repeating
as keys, then just key—what matters, the way out of labyrinth and code.
A chant or psalm, repeating
open vowels like water or rain. Luck—the river in poker, the silver ball
spun at the wheel’s rim—can be repeating
until it turns, and isn’t it possible to hear a persistent tune as anthem,
not ear-worm? Can’t repeating
strengthen a seam, or knit a long row? After you fell, I imagined you there
on the floor, the refrigerator’s hum repeating
its cold comfort until the ambulance came. Your husband cradling you
on the linoleum, repeating
presence in absence in presence in absence—cognitive dissonance—
or an inscrutable charm repeating
its mysteries. Your hip and arm are mostly healed. The house is still there,
but an interior door slams shut every second, an alarm repeating
its warning: all doors soon will be sealed. Where we stand in an earthquake
is an empty doorframe. Soon, repeating
itself will disappear: all language flown, a slaughtered lamb,
a shattered paradigm. Will your repeating
heart just—miss its cue? Will you forget how to swallow? O Fates, come, come,
/ Cut thread and thrum, repeating
its end-times refrain. Your jacks and pick-up sticks—you still love any game
—played now with your left arm, repeating
that makes of small movements an art, like dance. Your little jokes,
like the “ALZ tip jar”—now crammed—repeating
the ring of each coin dropped in. You pull my leg when I call, pretending
to forget who I am, repeating
Becky who? Becky who? until we both laugh, the sounds that sound so much
like crying but with a whetted edge; in time, repeating
a knife against stone will bring forth a terrible blade. O sweet sad funny sister,
you are the bird who flies away in my dream, repeating
its looping script on the blank page of sky. An unhooded hawk, a wild bird
freed. Calling Key? Key? Key?—repeating, repeating, repeating.
Source: Poetry (May 2021)
by way of: www.poetryfoundation.org/poetrymagazine/poems/155783/alz-ghazal
#poetry#ghazal poetry#Rebecca Foust#holy crap not a reblog even#the part where the poet compares cooking to jazz to sheets of empty musical staffs...!! 😵💀 I am slewn
2 notes
·
View notes
Text
Magiautotrophs
When Olua is deep into star-feeding, magic floods the atmosphere, and the air fills with critters who feed on it. It's so thick with life, certain macrofauna swim the sky throughout summernoon. Many microorganisms have developed ways essentially eat magic. These magisynthetic processes are fundamentally different from photo- or chemosynthetic ones, leading to a new set of trophic categories for Oluan microbes: Magiotrophs. An organism may get their energy, their electrons, or both from magic. An organism who does both and obtains carbon via CO2 fixation would be fully termed a magiomagiautotroph -- magiotroph for short. Though the convention is to shorten a -troph to its first (and sometimes third) prefix, a non-magisynthetic organism who nonetheless oxidizes magic for electrons will often be called a magitroph anyway, due to the novelty and contextual significance of the concept.
Side note
Though plenty of species use magic in myriad ways, including various energy-boosting strategies, no known eukaryote has figured out how to use magic directly as food, the way plants use sunlight. Magic, by definition, is poorly understood, so it is yet unknown why this is a power exclusive to the micros.
One popular hypothesis posits the ability originated in domain Alcana and spread via horizontal gene transfer to some other species; among alcans with fully sequenced genomes, over 90% have genes that code for magisynthetic biomachinery. Same and similar genes have been found in several bacteria and archaea (some of which are relative newcomers to Olua). Trace amounts of these genes have even been found in lichens. Though the magisynthetic section of archaea and bacteria is much smaller, that they possess these sequences at all implies remarkable things for interdominal HGT, and the role magic may play in the process.
Second side note
There is emerging evidence certain alcan lineages have evolved ways to avoid cleaving oxygen during carbon fixation, likely due to the sheer abundance of atmospheric oxygen and magiparticles' resemblance to it. Big if true.
(Is it possible some of these microbes have complexes unrelated to Rubisco at all?)
#worldbuilding#olua#biology#microorganisms#magic#unreality#that's right boiz olua's got midichlorians >:333#Proud of this one.
3 notes
·
View notes
Note
🔫 hand over the lore toxi
OK SO quick context: I was feverous last night, and when I’m feverous I semi-hallucinate Horrors.
This is gonna be LONG
Shadowknights are born when a mortal dies with a heart full of hate and betrayal. These souls are pulled down into the nether by Shad, and reborn as shadow souls.
shadow souls spend their first century in a converted Nether Fortress called The Nursery.
Gene is the head of the Nursery, in charge of… how should we say… re-educating the shadow souls, and guiding them through their new majick.
When the shadow souls make their first vessel— twist their magic into something tangible— they are graduated to the rank of Fledgeling.
as fledgelings, they receive intensive training, in majicks, combat, deception, etc etc and then sent out into the overworld for their first kill.
The First Kill is a very important milestone for every fledgeling— both culturally and metaphysically.
they are meant to hunt down their old leader, be it a lord/parent/etc. this destroys the ties to their past, permanently cleaving themselves to the ranks of the Shadow-knights.
But in a more real way, their First Kill is also their first true meal.
The Shadow Knights don’t eat human food, obviously, so what powers them?
…why, magic, of course! particularly, soul magic. They eat the dead. any soul will feed adequately, (though the more magic, the better) but the intensity of the relationship is significant.
When they return victorious from the Overworld, they can begin being sent on missions.
Laurence is an interesting case.
Unlike other shadowknights, Laurence doesn’t… die. or rather, he’s never separated from his mortal body. He isn’t alive. He isn’t dead, either.
#[my post]#my post#aphblr#aphmau#aphverse#minecraft diaries#mcd#[ asks ]#aphmau mcd#shadow knights#dante anon 🟦#lore
31 notes
·
View notes
Note
Do you have more iterator immune system headcanons? I would absolutely love to hear them >:) (or just immune system facts in general those are really cool)

inspectors definitely act on a more macro scale when it comes to defenses. there isn't a single cell i can compare them to because the roles of immune cells are very specific and inspectors are pretty general in how they act (poking around crevices, attacking, etc.) i know their design is very hydra-inspired, but they also look like dendritic cells to me.

these cells are best described as the "brains" of the immune system (except they dont have real thoughts and are stupid because they are cells but. moving on). these cells sample the surrounding fluid and if they identify a protein that is foreign, they are the first step in the adaptive immune response. they are technically part of the innate immune system, but they bridge the gap
iterators have a lot of microbes in them that need to stay alive. they probably defend themselves through one of the various immune systems they can have. i think the most interesting is one that a lot of bacteria have, CRISPR-CAS. this defense is primarily used against viruses. when viruses inject their genetic material into the bacteria, the bacteria will take a part of the virus's genome and add it to its own for memory (CRISPR). rna is then transcribed from this dna and cleaved into individual parts, each with one with one piece of rna from one virus. then, with tracer RNA (tracRNA), bind to the CAS9 protein and this complex searches the cell for the matching DNA sequence. when found, it will unwind the viral dna and chop it up, rendering it ineffective as more proteins and enzymes dissolve the DNA. yes this is where gene editing was discovered
this part is speculation, but maybe it helps with iterating. iirc new ideas are thought of when the microbes grow and change :p
19 notes
·
View notes
Note
Let's play a game of how much Lumian lore you can dump on me so I can compile a complete post and use it for world building for a Lumian character of mine (pls give me the knowledge I crave it)
Oh god. Ok you're on
I'll do my best 😵💫 if you're looking for anything else / more specific, lmk!
Keep in mind my bs can and will differ from everyone else because everyone is either normal about them or is also weird about them but this time with tails instead, and also in a fundamentally unique way from my shit. And I have a LOT.
So,
Some world setup, because it matters to me; KirinDave as a concept is like 3 characters - Kirin (Craftia), Quetzal (Earth), and Chimera (Twilight). There's other deific rulers of other places but they don't matter rn. Kirin is the god of the Heart, Chimera the Mind, and Quetzal the Form. The 3 have their own backstory but I'll skip it for now.
Quetz came in contact with their people, the humans, in early historic times they were completely terrified of them. Out of shame, Quetz invented different religions to interact with them in a way they could better understand, and chose forms that were more pleasing and comprehensible. Under these guises, these personas, Quetzal fell in love with several humans and they just kinda sired a bunch of demigod children as a result.
Kinda messed up but I'm choosing to cover it up and say the humans they loved knew what they were getting into when they agreed to have sex with a god, including the whole polyamory thing. For all intents and purposes everyone was cool with it except general society who took one look at these "horrific hybrid" half-human half-animalish demigod kids and freaked out, killing nearly half of them and their parents in the process.
This didn't happen like, all at once, just over time for a while until Quetz couldn't take it anymore and decided to relocate the remaining humans, their children, and a good chunk of people who were supportive and capable of running a society successfully, and had an arsenal of angels help relocate them to a planet so far away, Quetzal's Siblings couldn't detect them easily. They kept this planet so secret that they would've loved to take this secret to the ends of the universe and with their deaths if they could've, but you see...
In no time at all, this society came together and grew into one species; The Lumians.
Lumians did not retain their Origin story and have no idea where they came from. The Guardian Angels cleared their ancestor's minds of Earth, and they simply began anew, knowing only their relationships and skills. Lumians became a race of highly scientifically progressed people whose genetics are influenced by the "throw a dart at a board to see what animal you land on" genes of their demigod ancestors, and the naturally adaptive influence of being descended from the god of Life itself.
They were born and raised on a harsh planet with a huge sun and massive insectoid monsters, so they naturally thrived in caves and darker biomes, where they quickly evolved to glow. Soon, their bodies rapidly changed to include pointed ears with sharper hearing to better listen for specific bugs, sharper teeth to cleave through incredibly tough bug meat, stronger/bigger bodies and longer lifespans to survive with, and colored blood/glow for attraction reasons bc blood color is pretty meaningless to their society and it has little to no effect on their bodies or survival.
Lumians have a superiority complex to humans, and assume that they're the result of ancient Lumians who colonized Earth and "devolved". This is a point of contention in their society and anywhere else in space that Lumians interact with humans. Funny enough, despite being smart with technology and science and weaponry, Lumian society valued art and creativity so poorly that entertainment is virtually nonexistent here. They get their kicks by starting and joining wars across space, and partaking in what human culture they can capture from lightyears away.
Their buildings and cities are very sterile-looking, and what color they do intentionally use are organized in mathematical and artificial/generated ways. Idk how to explain it because I haven't looked for architecture that displays this, but think of pretentious billionaire houses. That's kinda what we're working with. Lumians also don't have a class system, so even a lowly outback-type reclusive farmer or woodsman with a small 2x2 home has access to the same rights, funding, and technology as everyone else.
Their planet, Lumina, has a large variety of biomes with jungles, swamps, deserts, temperate zones, rocky barrens, and so on. They only have one ocean, and it's about the size of South America, and in their southern hemisphere. Everywhere else are lakes, rivers, and an incredible amount of underground pocket water and plants that retain a lot of water (like "cut the root open and drink it like a straw" amounts). They do have some mammal-like creatures, but they are usually very tiny and serve as pets.
Some dogs came with their ancestors on the trip and they've also evolved! Due to the strange food they've fed on, they've gotten sleek coats, bigger and tougher teeth, pointy tongues, bigger/wider ears, and very unusual colors and eyes. I'll have to draw them later!
They have similar careers to us, but their society celebrates joining the armada, to an even bigger extent than the US. To them, it's THE job to have, the most noble position to take, you basically become a hero to your family and town. For all interns and purposes, they absolutely are a military state. The leader of the armada is Xephos's dad and he is basically a rockstar and democratically elected emperor of the planet. Matching that, their language is also monolith, but has distinct accents and dialects. I've written the alphabet somewhere on this blog in the past. They do have actual leaders of society that lead certain areas, very much like our countries do, but they all take a backseat to Xeph's father in the end. Everyone in general follows along with him in a similar fashion to how certain white people obsess over the royal British family.
Religion can be niche and taboo depending on the region, and magic is a marginal subculture in general. Lumians don't care a whole lot about bloodlines, lineages, family cultures, so on; everyone is motivated to become independent of themselves, and visiting family is socially accepted to be a rarity. Sometimes their bodies can have more animal-like features, like colored patterns, patches of floof, different shaped pupils or more "wild" irises, sharper nails, maybe even a tail who knows - but these physical differences are as rare as unique physical differences on Earth (i imagine white and black blood could even be possible if the Lumian were albino/melanistic). They have their own spaceships, news, radio, television, school systems, so on so forth. They just don't have a lot of variety in life and it kinda sucks. they suck lmao
Quetzal's secret is now as thinly veiled as tissue paper, thanks to the Lumians' exploration and interaction with Earth and Craftia. Humanoids are very distinctly localized to our solar system, and at least two of the deities are needed to create a living/sentient creature, so a humanoid race existing outside the solar system is definitely a huge cause for suspicion.
also Xeph's dad is planning a secret invasion of Earth and enslavement of humanity and he's overall a horrible person, but only in secret. to the public he's just the bee's fuckin knees.
I hope u like ;w; search my blog for other Lumian stuff, bc I've posted quite a bit about them! ALSO TAG ME IN ANY OCs.....I LIVE FOR THAT SHIT DUDE
25 notes
·
View notes
Note
❛ if you take side with my enemies , i will not hesitate to cleave your head from your shoulders. ❜

@saishuu-heiki

"cleave it then!"
Rem screams, pushing against that initial fear with the only other thing she knew: anger. It floods her veins, boiling through the hollow channels of her body and causing her limbs to shake. The rage courses through her, eyes filling with the cerulean glow of their shared genes, pupils thinning and her vision feels sharper. Her blazer is torn from her body, laying in a crumpled heap behind her as she storms forward, boots thudding against the concrete of the roof; brass knuckles gripped tightly in her right hand to the point of her joints screaming from the pressure of her grasp.
"tear it from my shoulders with your bare hands!"
rem shouts, that churning in her stomach causing waves of nausea to crest and fall like a wave. Her voice breaks, quivering with the unbridled rage she feels towards the image of his figure. He had to be real this time; the thrumming of her heart matched his own-- she felt his breath with every inhale of own lungs and her cells vibrate with intensity: they call to him, cry for a reunion between the two of them. She barely feels the tug at the center of her core: the lead that cinches tighter and tighter is ignored as she stands but a few feet from his towering body.
"They are my family.. they are my home."

she snarls, fangs gleaming as exasperation waivers her voice. She abandons the half-jargon she usually speaks, ire clarifying her words. Rem heaves, panting as she stands before the general in a sullied uniform; red dries to a rusty colour, staining the whites of her shirt and there are patches of crimson that are drying against the black slacks. The blood isn't hers, but of her comrades-- of the other turks and she feels tears prick at the corner of her eyes.
"we.. are not your enemies. you are your biggest adversary. your hatred blinds you and makes you a fool!"
her heart aches, the voices in her head writhing like smoke and screaming out for his touch. Her skin crawls, goosebumps breaking across the surface of her flesh. She's hot to the touch, outrage causing a flush to break across high cheekbones and her lip is tender from a wound that would bruise in the coming days. She feels a hiccup strangle her throat, and Rem tries to speak past it.
"if you are a man of your word then do it! end my suffering and bring this reunion she wants so desperately to happen, but know this, Sephiroth. The planet will not weep for you as it will for me!"
4 notes
·
View notes
Photo

Figure 4.4 The SREBP system. The full-length SREBP protein is located in the endoplasmic reticulum (ER, a system of membranous cavities within the cytoplasm). It is associated with the SREBP cleavage activating protein (SCAP), which “senses” the level of cholesterol, or related sterols, within the membrane of the ER. When the cellular cholesterol content is low, the SCAP-SREBP complex migrates to the Golgi complex (not shown), where specific proteases cleave SREBP to release the N-terminal portion, “mature” SREBP. Mature SREBP moves to the nucleus where it binds to sterol response elements in the promoter regions of many genes. If SREBP-2 is concerned (as shown in the figure), these are mainly genes concerned with cholesterol metabolism (LDL receptor, enzymes of cholesterol synthesis). SREBP-1 appears to be regulated more by expression of the full-length protein (which is increased by insulin); its proteolytic cleavage is also stimulated by insulin independently of cholesterol. SREBP-1c increases the expression of genes concerned with fat storage (including acetyl-CoA carboxylase and fatty acid synthase).
3 notes
·
View notes
Text
Philosophers have grappled with the nature of evil for thousands of years, but these days, immorality can feel like a solved problem. Take the case of Bryan Kohberger, the prime suspect in a quadruple homicide near the University of Idaho whose arrest ignited rampant media speculation about the psyche of a killer, as if properly diagnosing his personality disorder could mitigate the damage already done. His “psychopathic stare” made headlines in UK tabloids, while The New York Times dissected Kohberger’s self-described feelings of remorselessness as an adolescent. Dr. Drew brought on a former FBI agent to discuss Kohberger in the context of the “dark triad”: narcissism, psychopathy, and Machiavellianism.
Americans understandably want help making sense of the otherwise senseless deaths that populate the front pages of local papers and constitute Netflix’s extensive true-crime back catalog. But attempts to characterize evil remain scientifically dubious, say criminologist Jarkko Jalava and psychologist Stephanie Griffiths, coauthors of The Myth of the Born Criminal. When it comes to crime, psychologists frequently “get really sloppy,” Jalava says, adding, “we’re functioning on this folkloric level.”
The perpetrator of the University of Idaho murders should be condemned, but getting inside the mind of a killer is easier said than done. Prediction and prevention—the supposed end goal of criminal profiling—is even harder. And the proliferation of quasi-scientific terms for jerks, assholes, and even killers has far-reaching consequences.
The medicalization of evil—that is, the physician-led diagnosis and management of diseases like “moral insanity” and “criminal psychosis”—stretches back to the early 19th century. Where clerics once drew the line between good and evil, psychiatrists began to take people who engaged in impulsive, self-defeating, or otherwise un-Christian acts into their care.
Early on, these doctors-cum-criminal-profilers explained bad apples through theories such as atavism. Proponents believed that, over time, bad breeding led to degeneration of the gene pool, and the concentration of poverty, criminality, and other undesirable traits in certain ethnic groups or social classes. While the theory of degeneration was slowly replaced by a strikingly similar notion of “psychopathy” (literally “soul sickness”), many of the concerns remained the same: deviants who showed a lack of remorse or guilt, exhibited sexual promiscuity, and developed a lengthy rap sheet, perhaps from a young age.
New variations on this theme pop up all the time. The “dark triad,” coined in 2002 by Canadian psychologists Delroy Paulhaus and Kevin Williams, aims to describe “offensive but non-pathological personalities,” including CEOs, politicians, and bad boyfriends. There are also labels like antisocial personality disorder, a diagnosis given to individuals with severe impulsivity, aggression, and criminal behaviors—in other words, a DSM-approved twist on the old “psychopathic” standard.
At first glance, these attempts at categorization appear to be trending positive. For one thing, researchers are slowly cleaving obvious wrongdoing from the more inadvertent harms of mental illness. Similarly, it’s a relief to be able to use the dark triad to acknowledge just how commonplace selfishness really is.
But the shadow of degeneracy still looms large. In addition to further medicalizing everyday discourse (“jerks,” Jalava and Griffiths point out, have become “psychopaths,” with all the attendant baggage), these models uphold the dubious belief that every human has an immutable personality—and that those personalities can be easily classified as good or bad. In reality, recent research shows that many people change—and, in some cases, change dramatically—over the course of their lifespan. At the same, many researchers remain critical of the historic characterization of personality disorders, in part because it is stigmatizing and can obfuscate trauma, and even then it doesn’t lead to clear directions for treatment.
Many popular ideas about evildoers seem to stem from tabloid news, rather than scientific evidence. For example, Jalava and Griffiths have shown that many experiments that make claims of a genetic or neurobiological basis have not been replicated, and those that have been replicated have produced contradictory results. Soon, they will publish a new review detailing similar problems in studies of psychopathy and fMRI brain imaging. Perhaps most importantly, the husband-wife duo have documented how meta-analyses of psychopathy research, ostensibly the gold standard in scientific research, often ignore published results with null findings.
Even if researchers working on the problem of evil content themselves with the shortcomings of the existing scientific literature on psychopathy, dark triad, and the like, a new issue arises: People don’t just want to describe existing traits. They want to use these scales to predict future behavior.
In the criminal system, the results of assessments like the Hare Psychopathy Checklist are used to assess an individual’s risk of recidivism, and therefore the possibility or terms of parole. Paulhaus, the creator of the Dark Triad Personality Test, wants prospective employers, including police and the US military, to screen prospective candidates with his scale—and not always for the reasons you’d imagine. “It makes a big difference whether you diagnose someone as a Machiavellian or a sadist or a narcissist,” Paulhaus says. “There may be occasions where that's what you’re looking for.” At the same time, it’s easy to imagine a world in which individuals are screened for psychopathy and, if their score is high enough, closely monitored for potential crimes.
But Jalava and Griffiths insist that meaningful prediction isn’t possible with existing measures, either. If you remove questions on psychopathy evaluations that ask about previous criminal activity, these scales fail to anticipate what the test-taker does next. It’s an unsexy finding: “Past behavior can predict future behavior” is never going to be an A1 headline. But unlike other, more elaborate theories of pathological personalities, this at least is real.
The desire for strong language to match heinous acts is only natural. Unfortunately, sadistic bosses, everyday assholes, and even murderers are still human—shaped by and shaping the world. Condemning people as the subhuman (or, paradoxically, superhuman) embodiment of evil isn’t based on “the characteristics of the individual in front of us,” Griffith says, “but our response to them.” Such illusory categories make real understanding almost impossible.
In the context of evil, pursuing understanding is itself controversial. For decades, people have rightly criticized the media for the breathless attention it pays to killers—on the assumption that it feeds the psychopath’s or narcissist’s desire for attention, and may in turn inspire copycats. But there’s a difference between a news consumer’s insatiable desire for serial-killer content, and the responsibility we all have to face the harm in our society—and, perhaps, the capacity for harm in ourselves.
For example, many true-crime aficionados will cite data showing that something like one-third of serial killers (many of whom are presumed to be psychopaths) experienced physical abuse, one-fourth experienced sexual abuse, and half experienced psychological abuse as children. But these statistics don’t explain much at all about these “evil” adults, more than half of whom weren’t abused. Rather, they raise a more interesting question: Why are so many ostensibly non-psychopathic American parents abusive of their children?
This reformulation isn’t an excuse or an absolution; adults can, by and large, be considered responsible for their own actions. But it makes clear that “evil” doesn’t exist neatly in an individual in the way inherently judgemental labels like “psychopath” might imply. It’s not simply that psychopaths aren’t born but made, either. It’s that, if psychopaths exist at all, the same forces that shape them are at work on the rest of us, likely with similar, if more subtle consequences.
Flipping cruelty on its head also opens avenues for new solutions. We know, for example, that poverty is the number one cause of child abuse. One might reasonably conclude that the money devoted to studying dark traits might better be used for a universal basic income. Similarly, the notion that past behavior can, in meaningful ways, suggest future behavior is a helpful starting point. For example, if the only part of a psychopathy test that can predict crime are questions about past criminality, we should be able to ditch the armchair psychoanalysis and focus on documented, real-world behavior. Even then, because people can and do change, these rules cannot be hard and fast, and context and compassion will remain essential.
It’s still possible to divest even further from useless labels, by focusing on the idiosyncrasies of troubled—or, more precisely, troubling—individuals, as well as the specific context in which they emerged. Despite all of their scholarship, Jalava and Griffiths say they don’t have a silver bullet for defeating, let alone replacing, the myth of the born criminal. Rather, they advocate for a descriptive (rather than the standard normative) approach. Understanding how a certain person arrived at a certain point in time can’t undo the hurt they’ve caused, but it might help them to chart a new path—perhaps even one that deviates from their dark “personality.”
6 notes
·
View notes